2线粒体动力学
线粒体通过融合和分裂来不断地变化形态,这个过程称为线粒体动力学。线粒体融合和分裂之间的动态平衡是细胞功能正常发挥的重要条件。当线粒体动力学被破坏时,线粒体的形态和功能会受到影响,导致NAFLD的发生和发展。
2.1 线粒体融合
线粒体融合一般包括线粒体外膜融合和线粒体内膜融合两部分。线粒体外膜融合受到线粒体融合蛋白1/2(MFN1/2)的调控,线粒体内膜融合受到视神经萎缩蛋白1(OPA1)的调控。两个线粒体外膜上的MFN1/2形成同源二聚体或异源二聚体,并在GTP的作用下拉近两个线粒体距离,而后线粒体外膜开始融合。
2.2 线粒体分裂
参与线粒体分裂的蛋白主要包括线粒体动力相关蛋白1(DRP1)、线粒体分裂因子(MFF)、线粒体分裂蛋白(FIS1)、线粒体动力学蛋白49/51(MiD49/MiD51)。线粒体分裂位点通常在内质网与线粒体之间。细胞质内的DRP1受到线粒体表面的DRP1受体(MFF、FIS1、MiD49/MiD51)的募集后到达线粒体表面。DRP1被大量募集到线粒体表面,并与受体相结合形成环状结构后剪切线粒体,促进线粒体分裂过程。
3生物钟对线粒体动力学的影响
线粒体动力学受到生物钟的调控,表现出昼夜节律性变化。哺乳动物线粒体形态的变化表现出与外界光暗周期一致的昼夜节律性变化。培养的肝细胞中的线粒体表现出昼夜节律改变,线粒体的管状结构数量从亮到暗有节奏地减少。在肝脏中,BMAL1基因调控线粒体动力学基因的节律性表达,使其表现出与昼夜生物能量需求同步的代谢节律。研究表明,钙调磷酸酶的激活受生物钟调节,使DRP1在Ser637位点去磷酸化,促进DRP1易位到线粒体导致线粒体碎裂,Ser637磷酸化的DRP1水平表现出24小时节律。Xu等研究发现,CLOCK基因能够通过竞争性抑制PUF60功能来加速DRP1 mRNA的降解,PUF60可通过维持DRP1 mRNA稳定性直接调节线粒体稳态。昼夜节律紊乱时,线粒体融合和分裂会受到影响,当CLOCK基因丧失时可能会引起PUF60的释放,导致DRP1水平升高和线粒体碎片化。在肝脏BMAL1特异性消融的小鼠中,FIS1的基因和蛋白质水平的节律性被破坏,并伴线粒体肿胀和增大。
生物钟还通过一些介质调控线粒体动力学,例如Rheb、单磷酸腺苷激酶(AMPK)和sirtuins(SIRT)。哺乳动物雷帕霉素靶点复合物1(mTORC1)是线粒体活性和结构的主要调节因子。mTORC1通过促进线粒体分裂和生物发生来控制线粒体结构,促进受损线粒体自噬。这些mTORC1调节的线粒体事件表现出昼夜节律性,以响应代谢需求。大脑中富集的Ras同源物Rheb是体内mTORC1的重要激活剂。Yuan等研究证明,生物钟控制肝细胞中Rheb蛋白水平的节律性振荡,从而调节肝脏中mTORC1活性和DRP1稳定性。Rheb调控的mTORC1/DRP1轴有助于调节线粒体形态动力学,以实现最佳线粒体代谢。AMPK的活性、亚基组成和定位取决于生物钟。AMPK对线粒体动力学具有双重作用,AMPK可以在高能应激下通过磷酸化MFF触发线粒体分裂,有利于从细胞质中募集DRP1来提高线粒体分裂速率,促进线粒体自噬并迅速从线粒体自噬产物中获取能量。AMPK还可以在禁食期间通过控制MFN2基因促进线粒体融合,进而提高ATP生成率。生物钟还调控介导烟酰胺腺嘌呤二核苷酸(NAD)合成的烟酰胺磷酸核糖转移酶(NAMPT)的昼夜节律性振荡,引起NAD水平的昼夜节律性振荡。SIRT是NAD依赖性蛋白质脱乙酰酶,研究表明,生物钟可以通过NAD/SIRT3通路去乙酰化OPA1介导线粒体融合,SIRT3是线粒体中的一种重要的去乙酰化酶。综上所述,线粒体动力学受到生物钟的调控,并表现出昼夜节律(图1)。
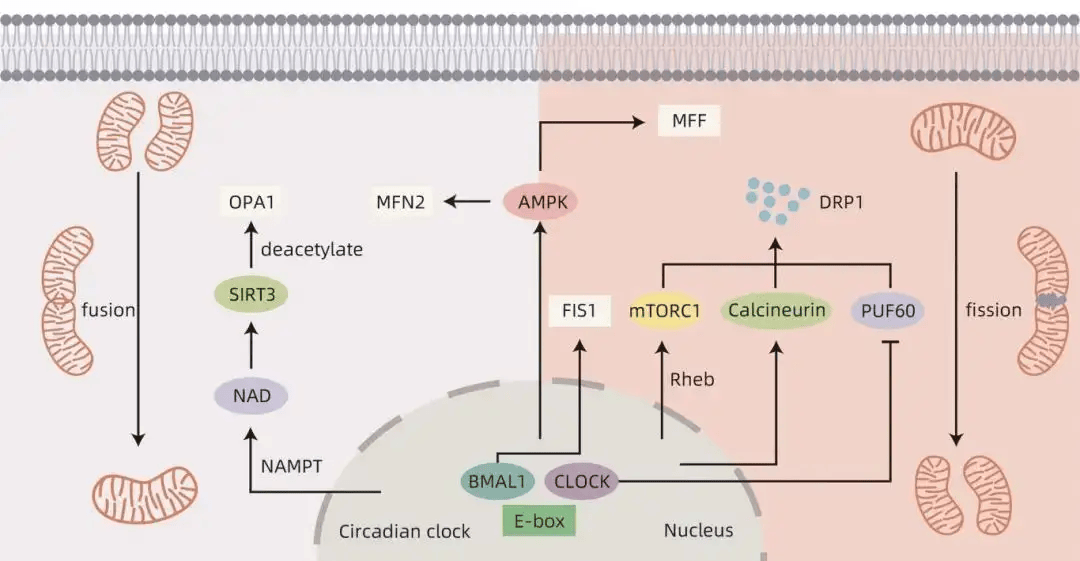
图1 生物钟调控线粒体动力学的相关机制示意图
4NAFLD中生物钟调控的线粒体动力学
熬夜和轮班工作等行为会扰乱人体内固有的、规律的生物钟,使机体内部生物钟与外界环境变化不同步,扰乱机体的生理和行为节律,引起昼夜节律紊乱。昼夜节律紊乱已被确定为NAFLD的常见危险因素。紊乱的生物钟调控线粒体动力学通过多种潜在机制损伤肝脏细胞,包括胰岛素抵抗、线粒体氧化应激、内质网应激和线粒体自噬失常等,导致NAFLD的发生和发展(图2)。
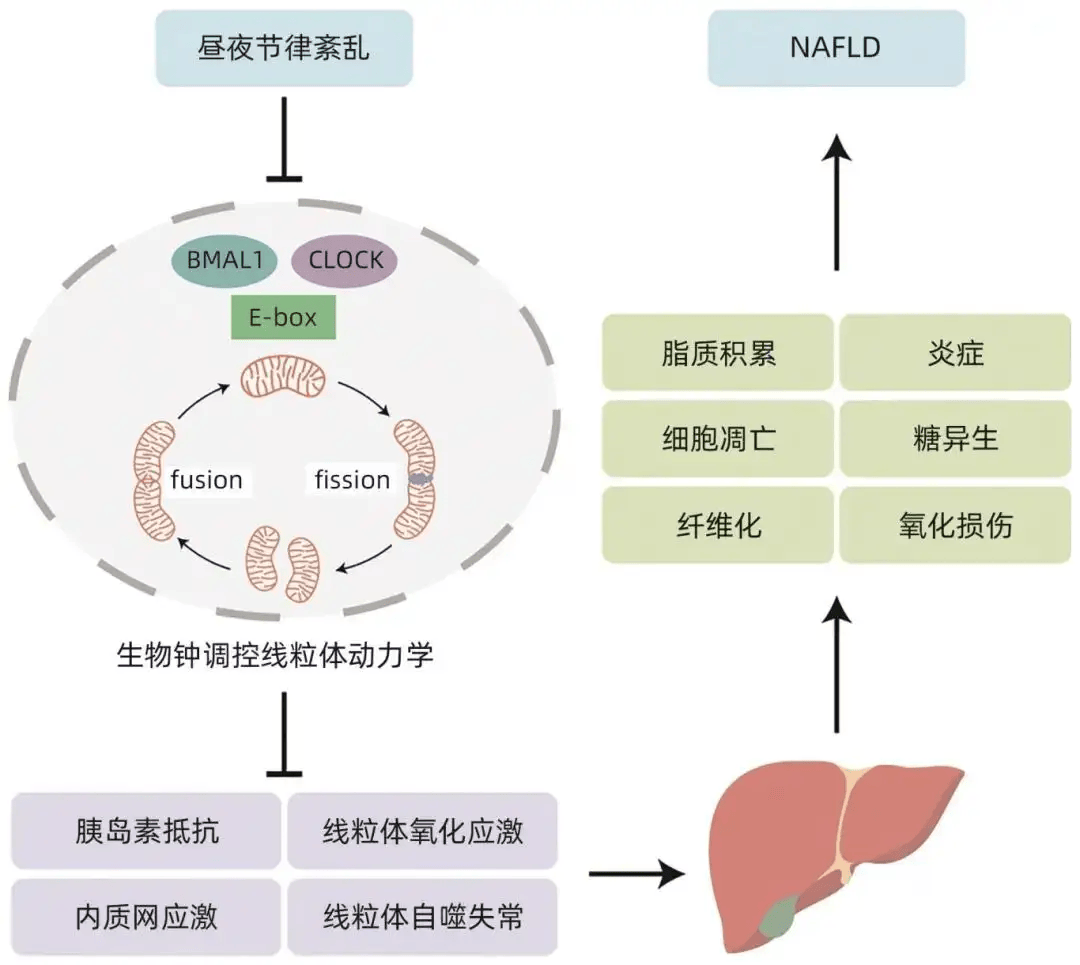
图2 生物钟调控的线粒体动力学在NAFLD中的作用机制示意图
4.1 生物钟调控的线粒体动力学和胰岛素抵抗
胰岛素抵抗时机体对葡萄糖摄取和利用率下降,血糖异常,机体为供能会促进脂质合成,抑制脂肪酸分解作用减弱,引起肝脏内甘油三酯聚集过多,超过肝脏的代谢和转运能力,导致肝脏内发生脂肪沉积,形成脂肪肝,是NAFLD发生发展的主要原因之一。胰岛素抵抗与线粒体动力学失衡密切相关。MFN1/2主要通过控制线粒体DNA的含量来调节葡萄糖刺激的胰岛素分泌,这对维持葡萄糖稳态至关重要。FIS1在调节胰腺β细胞代谢-分泌偶联中至关重要,在葡萄糖无反应细胞中FIS1的过表达增强了对葡萄糖的反应,增加胰岛素分泌并且使线粒体网络更均匀,在葡萄糖反应细胞中FIS1的下调减少了葡萄糖刺激的胰岛素分泌并导致线粒体网络不均匀。研究表明,INS-1细胞中的胰岛素分泌依赖于FIS1的表达,这一假设在原代小鼠β细胞中得到验证,其中FIS1的下调伴随着葡萄糖刺激的胰岛素分泌减少和线粒体网络不均匀。Kabra等发现β细胞特异性OPA1敲除小鼠表现出高血糖,β细胞增殖减少,ATP产生减少,胰岛素分泌减少。研究发现,线粒体动力学失衡会诱导氧化应激并停止产生能量,引起很多代谢信号通路异常,包括肝脏糖异生增强和炎症蛋白激活,长期的氧化状态和炎症会导致β细胞功能障碍和胰岛素抵抗。Mnf1/Mnf2的过表达和DRP1/FIS1的表达下调都能够通过激活胰岛素刺激的IRS1-Akt通路和诱导GLUT1/GLUT4易位到细胞膜来增强线粒体网络并改善胰岛素抵抗。综上所述,线粒体动力学通过影响胰岛素信号通路和胰岛素分泌,在胰岛素的生理功能中起着关键作用。
胰岛素抵抗也受到生物钟的调控。胰岛素的分泌表现出昼夜节律性变化,即白天分泌增加,夜间分泌减少,胰岛素受体在白天敏感性最高。昼夜节律紊乱可导致胰岛素抵抗的发生。长期轮班工作引起的昼夜节律紊乱是胰岛素抵抗的危险因素,轮班工作的工人出现胰岛素抵抗的概率更大。CLOCK基因突变动物表现出参与胰岛生长发育基因(CyclinD1、Gsk3β、Hnf4α、Hnf1α、Pdx1、NeuroD1)、胰岛素信号传导基因(InsR、Irs2、Pi3K p85、Akt2)和葡萄糖感应基因(Glut2、Gck)表达水平的下降和/或RNA振荡的相移。全身BMAL1基因敲除小鼠出现葡萄糖耐量降低、胰岛素分泌减少、胰岛素抵抗的表现。小鼠肝脏CLOCK/BMAL1基因敲减后,出现胰岛素抵抗,而通过尾静脉给糖尿病小鼠注射过表达CLOCK、BMAL1基因的腺病毒后,小鼠对胰岛素的敏感性得到改善。通过使用染色质免疫沉淀测序,研究人员发现CLOCK和BMAL1基因可以与胰岛素敏感性相关的线粒体内基因结合,这表明生物钟与胰岛素抵抗之间的关系可能与线粒体动力学有关。研究发现,BMAL1基因敲除小鼠既出现线粒体动力学失衡,又表现出了胰岛素抵抗。抑制BMAL1基因会导致线粒体动力学失衡,同时会损害胰腺β细胞中胰岛素的信号传导。因此,紊乱的生物钟可能通过调控线粒体动力学来引起胰岛素抵抗,并促进NAFLD的发生发展。
4.2 生物钟调控的线粒体动力学和线粒体氧化应激
氧化应激的特征是细胞内活性氧(ROS)水平升高或还原能力减弱而产生的不可控氧化过程。哺乳动物的线粒体是产生ROS的主要场所,氧化应激会损伤线粒体导致更多的ROS产生,如此形成恶性循环。线粒体功能障碍会导致大量ROS的生成,如果过量的ROS得不到及时清除,则会引起肝脏中线粒体和细胞中脂质和蛋白质的过氧化,进而促使线粒体的膜通透性转换孔开放,导致线粒体的膜电位降低甚至消失,破坏线粒体的结构与功能,造成肝脏功能的降低与损伤,在NAFLD的发生发展中具有关键作用。研究发现,NAFLD患者同时出现了氧化应激和线粒体功能障碍。相关研究报道了ROS水平和线粒体动力学的同步变化,这表明线粒体动力学与ROS产生之间存在内部关系。当代谢通量较大时,线粒体分裂增加与ROS水平升高同时存在,导致氧化损伤和细胞死亡。在游离脂肪酸混合物诱导HepG2细胞建立NAFLD体外模型中,线粒体动力学失衡,融合相关蛋白表达降低,分裂相关蛋白增高,同时伴有大量ROS产生,发生氧化应激。值得注意的是,在用高浓度葡萄糖或脂肪酸处理的细胞中,抑制线粒体分裂足以抑制ROS的增加和细胞死亡。因此,认为在代谢过量过程中,ROS增加可能是由线粒体分裂增加所致,并且可能是缓解与代谢疾病相关的氧化应激的靶标。后续研究发现,抑制线粒体分裂能够显著降低高脂饮食诱导的脂肪变性和氧化应激,这表明线粒体分裂在调节与NAFLD相关的肝脂质处理和氧化应激方面发挥作用,线粒体分裂是治疗NAFLD的新靶点。研究表明,DRP1激活导致线粒体分裂增强,随后ROS产生增加,而抑制DRP1导致ROS水平降低,在用高浓度葡萄糖或脂肪酸处理的细胞中抑制线粒体分裂足以抑制ROS增加和细胞死亡。线粒体分裂抑制剂-1(Mdivi-1)可以抑制细胞中DRP1的GTP酶活性,Mdivi-1可以通过抑制DRP1来逆转棕榈酸诱导的线粒体损伤和ROS产生。另一方面,ROS可能是DRP1激活的上游分子,过量的线粒体ROS引起DRP1过表达,引起线粒体分裂,出现线粒体碎裂增多。这些研究结果表明,线粒体动力学失衡和过量的ROS之间会相互作用,形成恶性循环,促进NAFLD的发生发展。
生物钟调控线粒体氧化应激的动态。线粒体和线粒体外ROS的产生,以及抗氧化系统状态呈现每日振荡。ROS包括羟自由基、超氧阴离子和过氧化氢(H 2O 2)。小鼠肝脏内源性H 2O 2水平在整个昼夜周期内也发生有节律的振荡。肝脏BMAL1基因缺失会产生过量的ROS,并导致脂肪肝和胰岛素抵抗。这些结果为ROS受到生物钟基因调控提供了有力证据。生物钟还调控抗氧化系统。核因子红系衍生物2(Nrf2)是一种经典的生物钟控制基因,其表达和激活表现出节律性模式,能够控制谷胱甘肽(GSH)的细胞内振荡水平并介导抗氧化反应。Aldh2是线粒体基因,具有抗氧化作用,其表达也受到生物钟的调控。受到生物钟调控的NAD +水平和SIRT3活性变化也调节线粒体氧化应激。Gong等研究发现禁食期间增加OPA1和减少Ppif表达可以提高呼吸效率,限制ROS的产生。相反,在饲喂期间降低OPA1和增加Ppif表达可促进ROS的产生。进一步研究表明,CLOCK基因可以通过稳定DRP1 mRNA表达来改善线粒体动力学和功能,在CLOCK基因突变小鼠中,DRP1的异常积累会导致线粒体片段化和ROS水平升高。由此可以推断,生物钟调控的线粒体动力学影响线粒体氧化应激,并在NAFLD的发生发展中起到重要作用。
4.3 生物钟调控的线粒体动力学和内质网应激
内质网主要负责蛋白质的合成、修饰和折叠,以及脂质的生物合成。细胞环境发生变化,如缺氧、ROS、营养缺乏等会引起内质网功能损害,导致内质网过量未折叠和/或错误折叠的蛋白积累,称为内质网应激。内质网应激会激活细胞未折叠蛋白反应(UPR),恢复内质网稳态或诱导细胞凋亡。肝细胞中持续的内质网应激会引起脂质代谢紊乱,最终UPR诱导细胞凋亡,引起肝损伤或肝细胞死亡,促进NAFLD的发生发展。哺乳动物的UPR响应主要涉及三条信号通路,包括蛋白激酶RNA样内质网激酶/转录活化因子4(PERK/ATF4)、1型ER转膜蛋白激酶α/X-box结合蛋白1(IRE1α/XBP1)和活化转录因子6(ATF6)通路。这三条通路都受到线粒体动力学的调控。MFN2介导线粒体与内质网之间的连接,参与线粒体相关内质网膜(MAM)的构建。研究表明,在MFN2缺陷细胞中出现UPR三条通路过度激活,促进内质网应激。此外,DRP1过表达会增加线粒体断裂和内质网应激,Mdivi-1能够通过抑制线粒体片段化来减弱内质网应激。
生物钟调控着很多内质网应激和UPR相关蛋白质的表达。PERK通路的中心转录因子ATF4受到CLOCK基因的转录调控,CLOCK基因可直接或间接影响PERK通路。IRE1α将UPR和生物钟联系在一起,小鼠肝脏中IRE1α/XBP1通路表现出生物钟依赖性节律激活,周期为12小时,这种节律还与BIP昼夜节律性表达有关,BIP是控制UPR激活的主要ER伴侣之一。研究发现,夜班工作导致昼夜节律紊乱的工人出现内质网应激相关基因的上调。生物钟的破坏扰乱了IRE1α介导的UPR激活的昼夜节律,并导致内质网酶的失调,这种破坏还导致甾醇调节元件结合蛋白(SREBP)转录因子的异常激活和肝脂质代谢受损,从而促进代谢紊乱的发展。此外,研究发现,昼夜节律紊乱引起内质网应激并影响UPR相关蛋白表达的同时伴有线粒体动力学失调。由此可以推测,恢复生物钟调控的线粒体动力学可能会抑制内质网应激,从而缓解NAFLD。
4.4 生物钟调控的线粒体动力学和线粒体自噬
线粒体自噬是一种清除受损线粒体的选择性自噬现象,能够调节细胞线粒体数量,维持细胞的正常功能。线粒体自噬主要的信号通路一种是PINK1/Parkin介导的线粒体自噬,另一种是PINK1/Parkin非依赖性途径,包括BNIP3、NIX和FUNDC1介导的线粒体自噬。线粒体自噬可以通过维持线粒体功能和蛋白质折叠过程来调节肝细胞死亡和脂质稳态。有证据表明,线粒体自噬异常会促进NAFLD的发生和发展。在NAFLD小鼠模型中线粒体自噬普遍减少,而增加线粒体自噬,可在去除损伤线粒体的同时减缓NAFLD进程。MFN1和MFN2的Parkin依赖性泛素化及其蛋白酶体和p97依赖性降解导致线粒体分裂增加,从而促进线粒体自噬。Xue等研究发现MFN2能降低BNIP3的表达,BNIP3介导的信号通路与MFN2控制慢加急性肝衰竭中细胞凋亡和自噬的平衡有关。此外,DRP1依赖性线粒体分裂可诱导线粒体自噬,DRP1能够使PINK1聚集和Parkin向线粒体易位,促进线粒体自噬。在成人心肌细胞中,DRP1是BNIP3诱导的线粒体自噬所必需的。过表达的FIS1会诱导线粒体片段化并促进形成自噬体包围线粒体,增强线粒体自噬。这些发现表明,线粒体动力学能够调节线粒体自噬。
线粒体自噬也受生物钟调控。小鼠肝脏中线粒体自噬的关键调节因子PINK1和BNIP3都出现昼夜节律性表达。小鼠肝脏中线粒体自噬是BMAL1基因的靶标,后者能够直接调节小鼠肝脏中线粒体自噬的介质(如PINK1和BNIP3)。BMAL1基因与BNIP3基因启动子中的E-box元件之一结合,BMAL1基因缺失会下调BNIP3蛋白水平,从而损害BNIP3介导的线粒体自噬。在肝脏中BMAL1特异性敲除的小鼠模型中,出现BNIP3的昼夜节律紊乱,线粒体自噬标志物的水平降低。此外,NR1D1(REV-ERBα)基因可以直接控制BNIP3介导的线粒体自噬,NR1D1基因敲除后出现BNIP3表达降低。Jacobi等研究表明,线粒体分裂和线粒体自噬蛋白在野生型(WT)小鼠的肝脏中表现出相似的昼夜模式。然而,在肝脏BMAL1特异性敲除小鼠中,它们的节律性被破坏,并且线粒体分裂和线粒体自噬相关基因 DRP1、FIS1、PINK1和BNIP3的表达也大大降低,表现出线粒体的形态或代谢缺陷,而FIS1的过表达可挽救肝脏BMAL1特异性敲除小鼠线粒体的这些缺陷。因此,线粒体自噬可能受到紊乱的生物钟调控的线粒体动力学的影响,从而促进NAFLD的发展。
5总结与展望
生物钟调控的线粒体动力学对肝脏的正常结构和功能至关重要。生物钟紊乱引起的线粒体动力学失衡会促进NAFLD的发生和发展。本文综述了生物钟调控线粒体动力学的相关途径,并讨论了紊乱的生物钟调控的线粒体动力学如何影响NAFLD的多种病因,包括胰岛素抵抗、线粒体氧化应激、内质网应激及线粒体自噬异常。本文为靶向调节昼夜节律和线粒体动力学来防治NAFLD提供了有力证据。目前对NAFLD的临床治疗,尚无有效的药物,主要采用非药物治疗干预手段,如运动、饮食控制和减肥手术等。未来可以将昼夜节律和线粒体动力学作为治疗NAFLD的切入点,更进一步研究生物钟调控线粒体动力学在NAFLD发生发展中的具体作用机制以及开展相关药物和治疗方法的研究。
全文下载 PDF & HTML
引证本文 Citation
张策, 苗嘉芮, 樊旭. 生物钟调控的线粒体动力学在非酒精性脂肪性肝病中的作用[J]. 临床肝胆病杂志, 2024, 40(8): 1670-1676
扩展阅读 Read more
特别关注|胰源性糖尿病的发病机制探析
特别关注|滤泡辅助性T淋巴细胞在自身免疫性肝炎中的作用机制
特别关注|不同来源的细胞外囊泡在肝细胞癌发生进展中的作用
特别关注|金属离子代谢:慢性肝病中医药防治新思路
特别关注|SREBP在非酒精性脂肪性肝病中的作用机制及治疗靶点
关于本刊 About us
■我国首个肝胆病专业杂志
■金色开放获取期刊(Golden OA)
■国际医学 期刊编辑委员会(ICMJE)规范期刊
■国际管理与技术编辑学会(ISMTE)会员期刊
■Embase、Scopus等13种国际重要数据库收录
■百种中国杰出学术期刊
■中国精品科技期刊
■中国百佳高校科技期刊
■收录期刊
■北京大学图书馆中文核心期刊
■中国科技论文统计源期刊
■RCCSE中国核心学术期刊
官方网站:lcgdbzz.org
官方微信:lcgdbzz1985
审稿专家申请入口:点击查看返回搜狐,查看更多
责任编辑:
平台声明:该文观点仅代表作者本人,搜狐号系信息发布平台,搜狐仅提供信息存储空间服务。
阅读 ()